A collaboration between

Crop Nitrogen Uptake and Partitioning
By Daniel Geisseler and William R. Horwath*
Javascript is disabled on your browser. For full functionality of this site it is necessary to enable JavaScript. Follow this link for instructions on how to enable JavaScript in your web browser.
Pattern of Seasonal Nitrogen Uptake - Overview
The cumulative N uptake curve of annual crops from planting to maturity generally follows an S-shaped (sigmoid) curve (see Figure). Little N is taken up during germination and seedling development. Nitrogen uptake is high and reaches a maximum during vegetative growth as the root system expands and leaves and stems develop. During the reproductive phase, the uptake pattern flattens off as N is moved or reallocated from the vegetative components to reproductive structures, such as seeds, fruits or tubers.
The period of rapid N uptake coincides with the time of rapid
biomass accumulation. In fact, when N supply is adequate, crop
N uptake during vegetative growth is mainly determined by crop
growth rate and is therefore affected by nutrient availability
and environmental conditions, such as water availability,
temperature and solar radiation. The N concentration in plant
tissue, such as leaves and petioles, strongly depends on N
availability. Tissue N concentrations can be used to determine
critical and optimal N concentrations in specific plant parts
at defined periods of growth. Timing and rates of fertilizer N
applications can be optimized based on critical N values to
increase N use efficiency and prevent N losses.
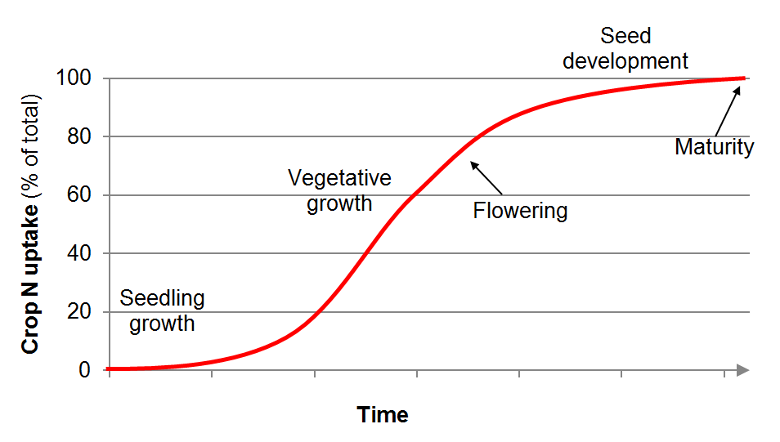
General shape of the N uptake curve for annual crops.
The N uptake rate does not always decrease before harvest. In crops that are harvested before flowers and seeds develop, such as lettuce and broccoli, N uptake remains high throughout the growing season. Nitrogen uptake may also remain high in crops with indeterminate growth. On indeterminate crops the flowers develop and open sequentially from the base towards the growing tip and the shoot continues to grow during early flower and fruit development. Examples are cotton or fresh-market tomatoes.
Nitrogen uptake in deciduous trees and vines also follows an S-shaped pattern between leaf-out and harvest. Nitrogen uptake is generally insignificant before leaf out. Most N needed in early spring is translocated from permanent storage organs, such as branches, trunk and roots. Nitrogen uptake is generally highest during leaf development and early fruit or nut development and slows during late development, as more N is translocated from the leaves to the fruits and nuts. Depending on the species, some N may be taken up after harvest to replenish N in permanent organs. When leaves senesce, N uptake from the soil ceases and part of the N in the leaves is remobilized and stored in permanent organs, where it is used the following spring to support early growth.
The N uptake curves shown for specific crops shall provide a general overview of the N accumulation pattern of these crops and are not intended to be used for detailed numeric fertilization recommendations.
Nitrogen Removed with Harvested Plant Parts
In many crops, only a proportion of the N taken up by the plants is removed with the harvested plant parts. The proportion of N removed from the field with harvested materials is crop-specific and may range in annual crops from about one third (e.g. broccoli) to almost 100% (e.g. silage corn or forage crops). The biomass left in the field will eventually decompose and part of the N will mineralize and become available to subsequent crops. What proportion of the N will be available for the next crop and how fast it will become available depends on the properties of the residue, such as its lignin content and carbon to N ratio, but also on soil, climate and crop management related factors that affect microbial decomposer activity. Furthermore, residual mineral N may be lost through leaching and denitrification between harvest and the establishment of the next crop as a result of precipitation and irrigation.
* Daniel Geisseler is an Extension Specialist in the Department of Land, Air and Water Resources at the University of California, Davis. William R. Horwath is professor of Soils and Biogeochemistry in the Department of Land, Air and Water Resources and the James G. Boswell Endowed Chair in Soil Science at the University of California, Davis.